Annals of Neurosciences, Volume 15, Issue 4 (October), 2008
Anurag Tripathi1, UC Srivastava2
1Département de zoologie, Ranchi College,(Ranchi University) Ranchi
2Département de zoologie, Université d’Allahabad, Allahabad, Inde – 211 002
Auteur corropondant
Dr UC Srivastava
Professeur de zoologie
Département de zoologie, Université d’Allahabad
Allahabad, Inde – 211 002
Téléphone : 93351 20178
E-mail : [email protected]
(Date de réception : 08.09.2008)
Abstract
L’acétylcholinestérase (AChE) met fin à la neurotransmission au niveau des synapses cholinergiques en scindant le neurotransmetteur acétylcholine. La nature et la distribution de l’enzyme ont été largement étudiées chez de nombreux invertébrés et vertébrés, y compris l’homme, par histochimie et biochimie. L’enzyme démontre un haut degré de variabilité dans la distribution avec sa présence notable dans les tissus non neuronaux également, ce qui fournit un thème pertinent pour étudier son rôle non classique.
Récemment, beaucoup d’informations sont sorties concernant sa structure dynamique, son expression génétique, son rôle dans la morphogenèse neuronale et la synaptogenèse. L’importance de l’AChE découle du fait qu’elle est la cible de médicaments conçus pour traiter la myasthénie, le glaucome, la maladie d’alzheimer, etc.
En gardant à l’esprit, les faits mentionnés ci-dessus, une revue approfondie a été faite dans le présent article concernant sa biochimie, sa dynamique structurelle, sa large distribution, ses isoformes et ses implications dans les troubles neurodégénératifs.
Mots clés : Acétylcholinestérase, cholinergique, formes moléculaires, troubles neurodégénératifs, maladie d’Alzheimer.
Introductions
L’acétylcholinestérase ( AChE) est l’une des enzymes les plus efficaces du système nerveux qui se concentre au niveau des synapses cholinergiques et des synapses neuromusculaires où elle hydrolyse rapidement le neurotransmetteur acétylcholine (Ach) en choline et acétate jouant ainsi un rôle essentiel dans la neurotransmission cholinergique.
Le terme acétylcholinestérase a été introduit en 1949 par Augustintion et Nachmansohn pour désigner une cholinestérase spécifique capable d’hydrolyser l’acétylcholine plus rapidement que les autres estérases. En 1964, la commission d’enzymologie a recommandé le nom « Acetylcholinesterase » (Acetylcholine Acetyl hydrolase ; 3.1.1.7) pour une cholinestérase véritable et spécifique.
La distribution de l’enzyme dans les tissus nerveux centraux et périphériques de différents vertébrés démontre une gamme élevée de variations (1-13). On a noté qu’elle était localisée dans les tissus non neuronaux et les cellules gliales également (14, 15). L’enzyme présente également une diversité moléculaire avec ses six formes moléculaires différentes et sa dynamique structurelle qui facilite son affinité et son action avec diverses légendes (16-17). En outre, on considère que l’AChE joue plusieurs rôles non classiques indépendants de sa fonction catalytique, à savoir l’hydrolyse de l’Ach. Ces rôles classiques et non classiques de l’AChE illustrent l’adéquation sur sa large présence dans les tissus neuronaux et non neuronaux (18-20).
L’importance de l’AChE dans l’homéostasie du corps est soulignée par le fait qu’ils sont les cibles de certaines des toxines les plus puissantes, y compris les insecticides, le venin de serpent et les armes chimiques (21).
Par conséquent, une telle large distribution et diverses fonctions, formes moléculaires, dynamique structurelle etc. de l’AChE fournissent une base adéquate pour rappeler qu’il s’agit d’une enzyme polyvalente, dont une connaissance détaillée, aidera à concevoir des médicaments spécifiques pour combattre diverses maladies neurodégénératives associées à cette enzyme.
Biochimie et dynamique structurelle
La structure tridimensionnelle de l’AChE a été déterminée pour la première fois en par Joel Sussman et.al en1991, en utilisant la protéine de T. californica (22). Les structures des domaines catalytiques des AChEs d’espèces comme T. californica, la souris et l’homme sont assez similaires (fig.1) (23, 24).
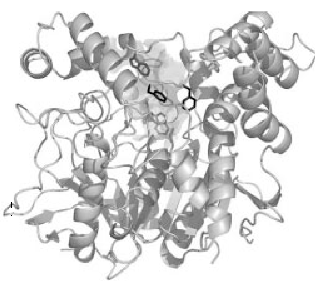
Figure 1. Schéma en ruban tridimensionnel de l’acétylcholinestérase de Torpedo Californica.
L’acétylcholinestérase est une sérine hydrolase appartenant à la famille des corboxylestérases de type B au sein des eucaryotes supérieurs. C’est une molécule ellipsoïdale d’environ 45× 60×65 A0, qui se compose d’un feuillet bêta mixte à 12 brins entouré de 14 hélices alpha. (22). C’est un monomère à l’état naturel avec un poids moléculaire d’environ 60 000 et forme des agrégats qui continuent à produire une activité catalytique.
Le site actif est composé de deux sous sites, le sous site estérique ayant la triade catalytique (Ser-His-Glu) et le sous site anionique de liaison périphérique (PAS) qui accueille le pôle quaternaire positif de l’Ach. ( Fig. 2a)
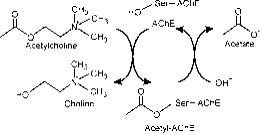
Figure 2a : Différents sites de l’AChE ; Le site catalytique est situé dans une gorge profonde.
Le sous-site estérique contient la machinerie catalytique de l’enzyme, une triade de Ser 200, His-440 et Glu-327 (les chiffres désignent la séquence des acides aminés dans la chaîne polypeptidique). Cette triade est similaire à d’autres sérine protéases, sauf qu’elle est la première à présenter Glu comme troisième membre, par opposition à Asp. En outre, la triade est de sens opposé à celle des autres protéases. Le sous-site anionique est défini par Trp 84, Phe 330, Phe 331. Son rôle est d’orienter la partie chargée du substrat qui entre dans le site actif. Cette fonction est principalement assurée par le Trp. 84 (22).
La gorge aromatique de la protéine a une profondeur d’environ 20 angströms et pénètre la moitié de l’enzyme. Le site actif se trouve à la base de cette gorge à seulement 4 angströms au-dessus de la base. La gorge aromatique est un terme plus approprié car 40% de son revêtement est composé de 14 résidus aromatiques qui sont hautement conservés de différentes espèces d’AChE (25).
Le site anionique périphérique a la capacité de se lier à de nombreux types de ligands différents par cela il affecte la confirmation du site actif. Six résidus stabilisent l’activité au sein du site. Trp 286, Ty r 72, Tyr 124, Glu 285, Asp 74, Tyr 341 situés sur le site opposé à l’entrée de la gorge. Cet ensemble de résidus offre une flexibilité qui permet d’accueillir de nombreux ligands et implique également leur mobilité de confirmation. L’AChE catalyse l’hydrolyse de l’AChE de la manière suivante (Fig. 2b):
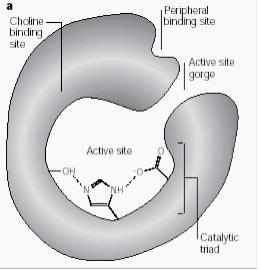
Figure 2b : Réaction du substrat de l’enzyme illustrant l’hydrolyse de l’acétylcholine.
La dynamique moléculaire de l’AChE est maintenue par un mécanisme de cross talk dans lequel une interaction entre deux sous-sites a lieu notamment entre les résidus Trp 286, Trp 86. Lorsque Trp 286 est lié à la périphérie, il affecte Trp 86 dans le site actif et provoque un site de confirmation distinct. L’Asp 74 joue également un rôle dans la modulation allostérique de l’enzyme. La sensibilité de ces résidus et la plasticité du site actif sont probablement le résultat d’une conception évolutive visant à conférer une activité optimale dans une grande variété de conditions (26.)
Occurrence
L’enzyme AChE est présente en forte concentration dans tous les types de tissus conducteurs, nerveux et musculaires, tissus centraux et périphériques, fibres motrices et sensorielles, fibres sympathiques et parasympathiques dites cholinergiques et non cholinergiques et toutes les régions où se trouvent les corps cellulaires et les jonctions, la concentration de l’enzyme est élevée(27). Avec les méthodes histochimiques, l’AChE est généralement mise en évidence dans les structures neuronales : périkarya, membranes de surface, axones, dendrites et régions synaptiques 4, 15, 28-30). L’activité de l’AChE est plus élevée dans les neurones moteurs que dans les neurones sensoriels (4, 15, 31). L’acétylcholinestérase dans les tissus nerveux est généralement reconnue comme représentant les neurones qui contiennent et ont vraisemblablement libéré l’acétylcholine en tant que neurotransmetteur (32).
Les premières études sur le site de synthèse de l’AChE dans le tissu nerveux ont indiqué une origine dans le corps cellulaire du neurone (33-35). On a constaté que l’enzyme était associée au réticulum endoplasmique du corps cellulaire et à l’appareil de Golgi (8, 36). L’AChE a également été associée dans les fractions subcellulaires du cerveau à des fractions microsomales et mitochondriales brutes (2, 37). Un fractionnement plus poussé de la fraction mitochondriale brute du tissu cérébral a montré des niveaux élevés d’activité AChE dans la fraction membranaire synaptosomale (6).
La majeure partie de l’AChE s’est avérée être localisée dans les axones et être associée à la membrane axonale (9). Une activité cholinestérasique limitée aux axones, a été observée sur les vésicules axonales de toutes les fibres nerveuses non myélinisées et de certaines fibres myélinisées (38). Les fibres nerveuses myélinisées ont en général une concentration plus faible que les fibres dites non myélinisées et la substance blanche contient beaucoup moins d’AChE que la substance grise (27). Dans les motoneurones isolés, l’activité de l’AChE est environ dix fois plus importante dans le cytoplasme, les dendrites et les axones que dans le nucléoplasme et aucune activité n’est présente dans le nucléole (39). Il a été démontré que les cellules gliales contiennent de l’AChE à des concentrations variables (14, 15)
Rôles classiques et non classiques
Tout le déroulement de la transmission cholinergique en particulier au niveau de la jonction neuromusculaire, à savoir la libération de l’Ach, sa diffusion à travers la fente synaptique, son interaction réversible avec le récepteur nicotinique de l’Ach et enfin son hydrolyse par l’AChE se produit en quelques millisecondes. L’ensemble du processus est étroitement intégré à la fois dans l’espace et dans le temps (40). Conformément à son rôle classique, son temps de retournement est de 150 usec, ce qui équivaut à l’hydrolyse de 5000 molécules d’Ach/molécule d’enzyme/seconde. Elle hydrolyse le substrat si rapidement que la concentration autour de la molécule d’enzyme est épuisée par rapport à sa concentration dans la solution en vrac.
Depuis plus de vingt ans, l’AChE est censée jouer d’autres rôles, en plus de son rôle classique de terminaison de la transmission synaptique. Ces fonctions non classiques de l’AChE pourraient impliquer l’hydrolyse de l’Ach dans un contexte non synaptique. Le peptide C-terminal ARP (peptide de lecture de l’acétylcholinestérase) de 26 résidus de l’AChE humaine, qui est produit lorsque le variant d’épissage R est induit par le stress, a été signalé comme modulant la différenciation hématopoïétique (41).
Diverses études in vivo et in vitero dans le système nerveux central ont suggéré que l’enzyme AChE régulée par le développement joue un rôle dans la fonction non cholinergique comme les processus morphométriques, la différenciation cellulaire et la synaptogenèse le long du système nerveux (20).De nombreuses études suggèrent que l’un des rôles non classiques de l’AChE pourrait être une protéine d’adhésion impliquée dans le développement et le maintien synaptique (42). Certaines autres évidences impliquent l’AChE en tant que protéine de la matrice osseuse, et il a été récemment démontré qu’elle interagit avec la laminine, protéine du sous-sol (43, 44).
Plusieurs études ont prouvé que l’AChE est impliquée dans la croissance des neurites. Des formes d’AChE hydrolysant l’Ach mais dépourvues des domaines neuritiques corboxy-terminaux (responsables des activités neuritogènes) n’ont pas réussi à favoriser la croissance neuritique, ce qui démontre l’indépendance de l’activité catylitique et neuritogène l’une par rapport à l’autre (19). On a également proposé que l’AChE joue de multiples rôles dans l’embryogenèse. Le poisson zèbre constitue un excellent système pour étudier les rôles invivo de l’AChE pendant l’embryogenèse. On a identifié une mutation par missence dans le gène ache du poisson zèbre qui abolit l’hydrolyse de l’Ach mais laisse intacte la partie corboxy-terminale neuritogène (45). On a également signalé que les embryons mutants ache présentaient des défauts dans le développement des fibres musculaires, la survie des neurones sensoriels primaires et la croissance dendritique. Ces défauts ont été collectivement interprétés pour soutenir des rôles non classiques de l’AChE.
Il existe de nombreuses preuves à l’appui de son rôle dans l’hydrolyse de la substance P, du met, de la leu-enképhaline et de la dégradation d’autres neuropeptides également (46, 47).
L’AChE a également été proposé pour fonctionner dans la morphogenèse cardiaque. L’activité AChE est détectée dans les cœurs embryonnaires de rat et de poulet avant l’innervation (48, 49). Bien que son rôle dans le cœur embryonnaire ne soit pas clair, il a été proposé que l’AChE régule un système de conduction des impulsions embryonnaires (50). Cependant, des fonctions supplémentaires sont nécessaires pour explorer davantage le rôle de l’AChE dans la fonction cardiaque.
Il a été démontré que l’AChE accélère l’assemblage du peptide αβ dans en fibrilles amyloïdes probablement par une interaction au niveau du site périphérique (51-53).Il a également été montré que les complexes AChE / αβ présentent une neurotoxicité accrue par rapport aux fibrilles ne contenant que des αβ (54).
Dans un contexte pharmacologique, les études ont révélé que les activités non classiques de l’AChE semblent converger vers le site anionique périphérique. On pourrait donc envisager que de nouvelles catégories d’inhibiteurs de l’AChE soient développées à l’avenir.
Diversité moléculaire
L’enzyme AChE se présente sous un ensemble de formes moléculaires différant à la fois par , la structure quaternaire et le mode d’ancrage au sein de la synapse générée par l’épissage alternatif de l’exon C terminal d’un seul gène, suivi d’une modification post-traductionnelle (55, 56). L’acétylcholinestérase (AChE) existe sous six formes différentes (isoenzymes) : monomère globulaire (G1), dimère (G2) et tétramère (G4) ; tétramère à queue (A4), tétramère à double queue (A8) et tétramère à triple queue (A12) (figure 3). La terminologie la plus rationnelle basée sur ces informations est celle développée par Bon et al. (1982). Dans leur système, les formes globulaires sont désignées par « G » et les formes à queue par « A ».
Les formes monomères et dimères à liaison disulfure peuvent être solubles ou attachées à une membrane par un glycophospholipide. Les formes tétramères peuvent être solubles, liées à une membrane par un lipide ou attachées à une triple hélice de collagène. Les formes monomères sont généralement incorporées dans les synapses du système nerveux central des vertébrés, tandis qu’au niveau des jonctions neuromusculaires, trois sous-unités catalytiques des tétramères sont attachées à une queue en triple hélice de collagène qui les ancre ensuite à la lame basale dans la fente synaptique.
L’expression des formes moléculaires de l’AChE, dans le Torpedo, a été observée comme étant spécifique au tissu suggérant que le polymorphisme présenté par l’AChE dépend d’un certain nombre de facteurs dont
la quantité d’Ach libérée
la taille de l’espace synaptique et/ou
le déroulement temporel de la stimulation physiologique.(58)
Régulation de l’expression de l’AChE dans les neurones
Ces dernières années, des études approfondies ont été menées pour élucider les mécanismes et voies moléculaires impliqués dans l’expression et la localisation de l’AChE dans les tissus neuronaux et non neuronaux. L’AChE est fortement exprimée pendant la différenciation neuronale. On a observé que la région non traduite 3′ (UTR) de l’AChE contient un élément riche en AU (ARE) qui interagit directement avec la protéine de liaison à l’ARN ‘HuD’, ce qui entraîne l’abondance des transcrits d’AChE dans les cellules neuronales (59). Ces résultats montrent l’importance des mécanismes post-transcriptionnels dans la régulation de l’expression de l’AChE dans les neurones en cours de différenciation et impliquent ‘HuD’ comme un facteur trans-actif clé dans ces événements. Cependant, plusieurs autres facteurs de régulation régissent la stabilité, la localisation et la traduction des transcrits de l’AChE. De nombreux éléments agissant en cis et facteurs et protéines agissant en trans interagissent avec l’AChE – 3′-UTR. (59).
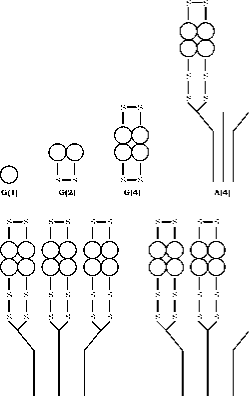
Figure 3. Structure quarternaire des six formes principales de l’AChE. Les formes globulaires sont représentées par G et les formes asymétriques par A selon le schéma de Bon et. al. (16). La localisation des ponts disulfures et le mode de liaison à la queue du collagène sont schématisés d’après le modèle construit par Anglister et Silman.(57)
L’AChE dans les troubles neurodégénératifs et neuromusculaires :
L’acétylchlinestérase a été directement liée à certains troubles neuromusculaires comme la myasthénie,le glaucome (60),et plus récemment pour pallier le déficit cholinergique associé à la maladie d’alzheimer (61).
Cliniquement, une inhibition modérée de l’AChE est efficace dans le traitementmen de ces maladies pour prolonger l’effet de l’ACh sur le récepteur Un tel traitement est souhaitable soit si la concentration d’ACh est réduite, comme dans le cas de la maladie d’Alzheimer, soit si les récepteurs sont moins nombreux, comme dans le cas de la myasthénie grave. Les inhibiteurs actuellement approuvés par la FDA américaine pour le traitement de la maladie d’Alzheimer sont l’E2020 (donépézil, Aricept), la tacrine (Cognex), la rivastigmine (Exelon) et la galantamine (Reminyl)(62). Les inhibiteurs utilisés dans le traitement de la myasthénie grave comprennent la pyridostigmine (Mestinon) et la néostigmine (Prostigmine)(63). Cependant, une inhibition trop importante de l’AChE, notamment par liaison covalente à la sérine du site actif, est invariablement mortelle. Par conséquent, l’AChE est une cible de choix pour les toxines naturelles telles que la fasciculine II du venin de serpent, les pesticides tels que le parathion et le malathion, et les agents de guerre chimique tels que le sarin, le tabun et le VX (64)
Conclusions
Dans le présent article, nous avons d’abord passé en revue les résultats récents liés à la biochimie et à la structure moléculaire de l’AChE qui fournit l’enzyme unique, la flexibilité pour rencontrer diverses légendes et aussi pour présenter un taux de rotation élevé. Nous avons également illustré son large schéma de distribution histochimique dans les tissus neuronaux et non neuronaux qui peut être corrélé à ses rôles classiques et non classiques qui ont été examinés. Des études supplémentaires sur la régulation de l’expression de l’AChE, qui révèlent l’importance de ces événements post-transcriptionnels in vivo, sont de la plus haute importance car elles pourraient conduire à la conception de stratégies thérapeutiques supplémentaires visant à promouvoir la régénération et la survie des neurones. Les développements récents dans la biologie de l’AChE, en particulier ses implications dans certains troubles neurodégénératifs, sont d’un intérêt capital et ont été brièvement passés en revue. Ces recherches aboutissent à la production de nouvelles classes d’inhibiteurs de l’AChE ciblées contre ses PAS (sites anioniques périphériques), qui pourraient jouer un double rôle dans le contexte de la maladie d’Alzheimer en inhibant simultanément l’hydrolyse de l’acétylcholine et en retardant l’assemblage du peptideá â en fibre amyloïde.
Abréviations
AChE – acétylcholinestérase ; ARP – peptide de lecture de l’acétylcholinestérase ; Ach – acétylcholine ; UTR – région non traduite ; ARE – élément riche en AU ; PAS – site anionique périphérique ; Glu – acide glutamique ; His – histidine ; Ser – sérine ; Asp – acide aspartique ; Trp – tryptophane ; Tyr – tyrosine ; Phe – phényl alanine.