1. Symétrie et taille
Les annélides sont tous des animaux à symétrie bilatérale. Leur taille varie de beaucoup moins de 1 mm de longueur à plus de 3 m. Les plus petits annélides décrits à ce jour sembleraient être Neotenotrocha (Dorvilleidae, Eunicida) atteignant seulement quelques centaines de microns de longueur (Eibye-Jacobsen et Kristensen, 1994). Les plus grands annélides se trouvent également dans la catégorie des Eunicida. Un spécimen conservé d’Eunice (Eunicidae) au Musée d’histoire naturelle de Washington D. C. mesure près de 3 m de long et environ 5 cm de diamètre dans son état conservé et contracté. Certains vers de terre, comme le ver de terre géant du Gippsland (Megascolides australis), peuvent atteindre 1 m de long. La plus grande sangsue du monde, Haementeria ghilianii, se trouve en Amérique du Sud et atteint plus de 40 cm de longueur.
2. Coelome
Presque tous les annélides ont une cavité remplie de liquide entre la paroi externe du corps et l’intestin, et c’est ce qu’on appelle un coelome (figure 1). Le cœlome est souvent utilisé comme zone de stockage des gamètes et sert de squelette hydrostatique pour la locomotion. Le cœlome (formé par schizocoïèse) est généralement organisé en une série de compartiments divisés par des septa intersegmentaires. Cette compartimentation signifie que si le ver est endommagé, le contenu coelomique ne sera perdu que dans quelques segments, et la locomotion peut être maintenue. Chez d’autres annélides, le coelome peut n’être divisé que par quelques septa. Dans ces circonstances, la perte de liquide coelomique est beaucoup plus importante en cas de blessure, et le ver peut être gravement affecté. Un certain nombre de groupes, notamment les sangsues et un certain nombre de petits polychètes, ont peu ou pas d’espace coelomique.

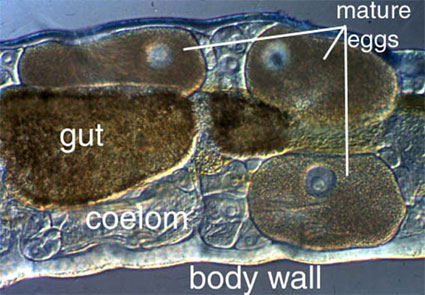
Figure 1. Amphicorina mobilis (Sabellidae). Bondi, Australie. Copyright © 1996 Greg Rouse.
3. Paroi du corps
Les annélides ont un corps recouvert d’une cuticule externe qui n’est jamais perdue ou mue. Les microvillosités épidermiques sécrètent un réseau de fibres qui sont en partie collagéniques et contiennent également de la scléroprotéine. Les chaetae sont également des structures cuticulaires, mais contiennent de grandes quantités de chitine. Sous l’épiderme et sa lame basale se trouve une couche de muscle circulaire. La couche de muscle circulaire forme une gaine presque continue autour du corps, sauf chez les polychètes dont les parapodes sont bien développés. Sous la couche de muscles circulaires se trouvent d’épais muscles longitudinaux. Chez de nombreux annélides, les muscles longitudinaux sont présents sous forme de quatre bandes distinctes (figure 2).

Figure 2. capricorne (Hesionidae) (Pleijel et Rouse, 2000). Île One Tree, Australie. Copyright © 2000 Greg Rouse.
4. Parapodes
Les parapodes sont des extensions segmentaires non jointives de la paroi corporelle que l’on trouve chez de nombreux polychètes, bien que beaucoup en soient dépourvus. Ils sont également absents chez les Clitellata et les Echiura. Les parapodes sont équipés d’une musculature dérivée principalement de la couche musculaire circulaire et portent généralement des chaetae. La structure des parapodes varie mais on peut considérer qu’ils se composent essentiellement de deux éléments : un notopode dorsal et un neuropode ventral (Fig. 3). En plus des faisceaux de chaetae, les notopodes et les neuropodes peuvent aussi avoir une variété de cirres et de branchies. Ils sont plus élaborés chez les formes rampantes ou nageuses actives où ils forment de grands lobes charnus qui agissent comme des pagaies. Les parapodes des polychètes fouisseurs ou tubicoles peuvent simplement être des crêtes légèrement surélevées portant des chaetae crochus appelés uncini.

Figure 3. Ophiodromus flexuosus (Hesionidae). Banyuls, France. Copyright © Fredrik Pleijel.
5. Système nerveux
Les anélidés possèdent un cerveau ou ganglion cérébral qui prend naissance et réside généralement dans la tête. La structure du cerveau varie, les formes actives mobiles ayant les cerveaux les plus complexes, et les formes sessiles ou fouisseuses ayant des cerveaux simples et peu différenciés. Il est relié au cordon nerveux ventral par les connecteurs circumpharyngiens, qui descendent de chaque côté du pharynx. Le cordon nerveux ventral, généralement constitué d’une paire de cordons reliés entre eux, s’étend sur toute la longueur du corps (Fig. 4). Il varie en épaisseur et se dilate en un ganglion dans chaque segment, à partir duquel des paires de nerfs segmentaires partent vers la paroi du corps, les muscles et l’intestin.
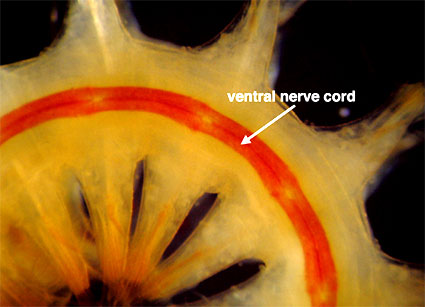
Figure 4. Vue ventrale d’un ver à écailles (Sigalionidae) montrant sa double corde nerveuse. Ce ver est inhabituel car il a une pigmentation rouge associée à la corde, ce qui le rend facilement visible. Misaki, Japon. Copyright © 1998 Greg Rouse.
6. Organes sensoriels
On trouve six grands types de structures sensorielles chez les annélides. Il s’agit des palpes, des antennes, des yeux, des statocystes, des organes nucaux et des organes latéraux (figure 5). Les palpes et les antennes sont situés sur la tête de nombreux polychètes. Chez certains groupes, ils sont tous deux sensoriels alors que chez d’autres, les palpes sont utilisés pour se nourrir. Les organes nucaux sont des structures chimiosensorielles ciliées, appariées, innervées à partir de la partie postérieure du cerveau. Ils sont présents chez presque tous les polychètes, et Rouse et Fauchald (1997) ont suggéré qu’ils pourraient représenter une apomorphie pour les Polychaeta. Ceci a été contesté par d’autres auteurs qui suggèrent que les organes nucaux pourraient être une apomorphie pour les Annélides dans leur ensemble et qu’ils ont été perdus chez les Clitellata/Oligochaeta (Purschke et al., 2000). Les Annélides possèdent également une variété de cellules sensorielles épidermiques qui peuvent être sensibles à la lumière ou au toucher (comme les organes latéraux).

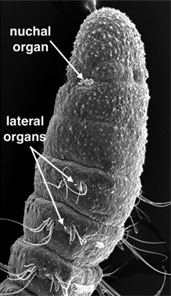
Figure 5. A gauche, Eumida (Phyllodocidae). Bundeena, Australie. Copyright © 1999 Greg Rouse. A droite, Proscoloplos (Orbiniidae). Bondi, Australie. Copyright © 2001 Greg Rouse.
7. Circulation et structures respiratoires
Un système circulatoire fermé est présent chez la plupart des polychètes (Fig. 6), les échiures et de nombreux clitellés. Il est réduit ou absent chez les sangsues où il peut être remplacé par des canaux coelomiques. Un système circulatoire limité, dans lequel certains des principaux vaisseaux sanguins sont présents mais les vaisseaux capillaires distaux sont absents, est présent dans un certain nombre de groupes de polychètes. Un système circulatoire est absent chez de nombreux petits polychètes.

Figure 6. Australonereis ehlersi (Nereididae). Bundeena, Australie. Copyright © 1999 Greg Rouse.
8. Organes segmentaires
Dans la plupart des annélides, il y a généralement deux systèmes fluides, le cœlome et le système circulatoire, et tous deux (s’ils sont présents) sont impliqués dans l’excrétion des déchets. Pour réaliser cette excrétion, il doit y avoir des conduits vers l’extérieur, et ceux-ci sont généralement appelés néphridies. Des conduits, appelés gonoductes ou coelomoductes, sont également nécessaires pour le transfert des gamètes qui se développent dans le coelome vers l’extérieur du corps à maturité. Les deux différents types de conduits sont souvent simplement appelés organes segmentaires, car déterminer quel type de conduit est présent est problématique.
.