Annals of Neurosciences, Volume 15, Issue 4 (October), 2008
Anurag Tripathi1, UC Srivastava2
1Department of Zoology, Ranchi College,(Ranchi University) Ranchi
2Departartment of Zoology, Universiteit van Allahabad, Allahabad, India – 211 002
Corresponderende auteur
Dr UC Srivastava
Professor in de Zoölogie
Departement of Zoology, Universiteit van Allahabad
Allahabad, India – 211 002
Phone : 93351 20178
E-mail : [email protected]
(Datum ontvangst: 08.09.2008)
Abstract
Acetylcholinesterase (AChE) beëindigt de neurotransmissie bij cholinerge synapsen door de neurotransmitter acetylcholine te splitsen. De aard en de verdeling van het enzym is uitgebreid bestudeerd bij vele ongewervelde en gewervelde dieren, waaronder de mens, zowel histochemisch als biochemisch. Het enzym vertoont een hoge mate van variabiliteit in distributie met zijn opmerkelijke aanwezigheid ook in niet-neuronale weefsels, hetgeen pertinente thema’s oplevert om zijn niet-klassieke rol te onderzoeken.
Recentelijk is veel informatie naar buiten gekomen over zijn dynamische structuur, genexpressie, zijn rol in neuronale morfogenese en synaptogenese. Het belang van AChE komt voort uit het feit dat het het doelwit is van geneesmiddelen die zijn ontworpen voor de behandeling van myasthenia gravis, glaucoom, de ziekte van Alzheimer etc.
Met het oog op bovengenoemde feiten is in dit artikel een grondig overzicht gemaakt van zijn biochemie, structurele dynamiek, brede distributie, isovormen en zijn implicaties in neurodegeneratieve aandoeningen.
Key words : Acetylcholinesterase, Cholinergisch, Moleculaire vormen, Neurodegeneratieve aandoeningen, Ziekte van Alzheimer.
Inleiding
Acetylcholinesterase ( AChE) is een van de meest efficiënte enzymen van het zenuwstelsel dat geconcentreerd is bij de cholinerge synapsen en bij neuromusculaire synapsen waar het snel de neurotransmitter acetylcholine (Ach) hydrolyseert in choline en acetaat en zo een essentiële rol speelt in de cholinerge neurotransmissie.
De term acetylcholinesterase werd in 1949 geïntroduceerd door Augustintion en Nachmansohn voor een specifiek cholinesterase dat in staat is acetylcholine sneller te hydrolyseren dan andere esterasen. In 1964 heeft de commissie voor enzymologie de naam “Acetylcholinesterase” (Acetylcholine Acetylhydrolase; 3.1.1.7) aanbevolen voor een echt en specifiek cholinesterase.
De distributie van het enzym in de centrale en perifere zenuwweefsels van verschillende gewervelde dieren vertoont een grote variatie (1-13). Er is vastgesteld dat het ook gelokaliseerd is in niet-neuronale weefsels en gliacellen (14, 15). Het enzym vertoont ook moleculaire diversiteit met zijn zes verschillende moleculaire vormen en structurele dynamiek die zijn affiniteit en werking met verschillende legenden vergemakkelijkt (16-17). Bovendien wordt AChE geacht verschillende niet-klassieke rollen te spelen, onafhankelijk van zijn katalytische functie, d.w.z. de hydrolyse van Ach. Deze klassieke en niet klassieke rollen van AChE illustreren adequaat zijn brede voorkomen in neuronale en niet neuronale weefsels (18-20).
Het belang van AChE in de homeostase van het lichaam wordt onderstreept door het feit dat zij het doelwit zijn van enkele van de meest krachtige toxinen, waaronder insecticiden, slangengif en chemische wapens (21).
Daarom biedt zo’n brede verspreiding en verschillende functies, moleculaire vormen, structurele dynamiek enz. van AChE voldoende basis om het een veelzijdig enzym te noemen, waarvan een gedetailleerde kennis zal helpen bij het ontwerpen van specifieke geneesmiddelen ter bestrijding van verschillende neurodegeneratieve ziekten die met dit enzym in verband worden gebracht.
Biochemie en structurele dynamica
De driedimensionale structuur van AChE werd voor het eerst bepaald in door Joel Sussman et.al. in 1991, met behulp van het eiwit van T. californica (22). De structuren van de katalytische domeinen van AChE’s van soorten als T. californica, muis en mens lijken sterk op elkaar (fig.1) (23, 24).
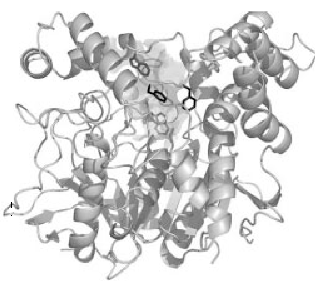
Figuur 1. Driedimensionaal lintdiagram van Torpedo Californica acetylcholinesterase.
Acetylcholinesterase is een serinehydrolase dat behoort tot de familie van type B corboxylesterasen binnen hogere eukaryoten. Het is een ellipsoïdaal molecuul van ongeveer 45× 60×65 A0, dat bestaat uit 12-strengs gemengde betavellen omgeven door 14 alfa-helixen. (22). Het is een monomeer in zijn natuurlijke staat met een molecuulgewicht van ongeveer 60.000 en vormt aggregaten die katalytische activiteit blijven voortbrengen.
De actieve site bestaat uit twee subsites, esterische subsite met katalytische triade (Ser-His-Glu) en perifere bindende anionische subsite (PAS) die plaats biedt aan de positieve quaternaire pool van Ach. ( Fig. 2a)
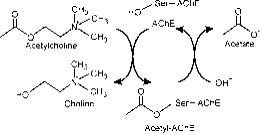
Figuur 2a: Verschillende sites van AChE; de katalytische site bevindt zich in een diepe kloof.
De esterische subsite bevat de katalytische machinerie van het enzym, een triade van Ser 200, His-440 en Glu-327 (de nummers geven de volgorde van de aminozuren in de polypeptideketen aan). Deze triade is vergelijkbaar met andere serine-proteasen, behalve dat de triade als eerste Glu als derde lid heeft in plaats van Asp. Bovendien is de triade tegengesteld aan die van de andere proteasen. De anionische subsite wordt bepaald door Trp 84, Phe 330, Phe 331. Zijn rol is het oriënteren van het geladen deel van het substraat dat de actieve site binnenkomt. Dit wordt voornamelijk gedaan door Trp. 84 (22).
De aromatische kloof in het eiwit is ongeveer 20 angstrom diep en dringt tot halverwege het enzym door. De actieve plaats ligt aan de basis van deze kloof slechts 4 angstrom boven de basis. Aromatische kloof is een meer passende term omdat 40% van de bekleding bestaat uit 14 aromatische residuen die in hoge mate geconserveerd zijn bij verschillende soorten AChE (25).
Perifere anionische site heeft de mogelijkheid om zich te binden aan veel verschillende soorten liganden, waardoor het de bevestiging van de actieve site beïnvloedt. Zes residuen stabiliseren de activiteit binnen de site. Trp 286, Ty r 72, Tyr 124, Glu 285, Asp 74, Tyr 341 gelegen op de tegenovergestelde site van de kloof ingang. Deze array van residuen biedt flexibiliteit die plaats biedt aan vele liganden en impliceert ook hun bevestigingsmobiliteit. AChE katalyseert de hydrolyse van AChE op de volgende wijze (fig. 2b):
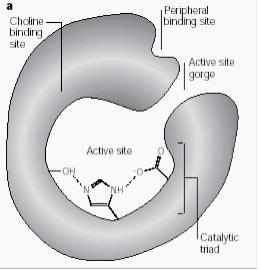
Figuur 2b: Enzymsubstraatreactie ter illustratie van de hydrolyse van acetylcholine.
De moleculaire dynamiek van AChE wordt in stand gehouden door het “cross talk”-mechanisme, waarbij interactie tussen twee subsites plaatsvindt, met name tussen Trp 286, Trp 86 residuen. Wanneer Trp 286 aan de periferie is gebonden, beïnvloedt dit Trp 86 in de actieve site en veroorzaakt het een aparte bevestigingsplaats. Asp 74 speelt ook een rol in de allosterische modulatie van het enzym. De gevoeligheid van deze residuen en de plasticiteit van de actieve site zijn waarschijnlijk het resultaat van een evolutionair ontwerp dat gericht is op het verlenen van optimale activiteit onder een grote verscheidenheid van omstandigheden (26.)
Occurrence
AChE-enzym is in hoge concentratie aanwezig in alle soorten geleidend weefsel, zenuw en spier, centrale en perifere weefsels, motorische en sensorische vezels, sympatische en parasympathische zgn. cholinerge en niet-cholinerge vezels en in alle gebieden waar zich cellichamen en knooppunten bevinden, is de concentratie van het enzym hoog(27). Met histochemische methoden wordt AChE in het algemeen aangetoond in neuronale structuren: perikarya, oppervlaktemembranen, axonen, dendrieten en synaptische gebieden 4, 15, 28-30). De activiteit van AChE is hoger in motorische neuronen dan in sensorische neuronen (4, 15, 31). Acetylcholinesterase in zenuwweefsels wordt algemeen erkend voor neuronen die acetylcholine als neurotransmitter bevatten en vermoedelijk vrijgeven (32).
Eerste studies naar de plaats van synthese van AChE binnen zenuwweefsel wezen op een oorsprong binnen het cellichaam van het neuron (33-35). Het enzym bleek geassocieerd te zijn met het endoplasmatisch reticulum van het cellichaam en het Golgi-apparaat (8, 36). AChE is in subcellulaire fracties van de hersenen ook in verband gebracht met microsomale en ruwe mitochondriale fracties (2, 37). Verdere fractionering van de ruwe mitochondriale fractie van hersenweefsel toonde hoge niveaus van AChE activiteit in de synaptosomale membraanfractie (6).
Het grootste deel van AChE bleek gelokaliseerd te zijn in de axonen en geassocieerd te zijn met de axonale membraan (9). Cholinesterase activiteit beperkt tot axonen, is waargenomen op axonale blaasjes van alle ongemyeliniseerde en sommige gemyeliniseerde zenuwvezels (38). Gemyeliniseerde zenuwvezels hebben in het algemeen een lagere concentratie dan de zgn. ongemyeliniseerde en de witte stof bevat veel minder AChE dan de grijze stof (27). In geïsoleerde motoneuronen is de AChE-activiteit in het cytoplasma, de dendrieten en axonen ongeveer tien keer zo groot als in het nucleoplasma en in de nucleolus is geen activiteit aanwezig (39). Van gliacellen is aangetoond dat zij AChE in wisselende concentraties bevatten (14, 15)
Klassieke en niet-klassieke rollen
Het gehele verloop van cholinerge transmissie, met name bij de neuromusculaire junctie, namelijk het vrijkomen van Ach, de verspreiding daarvan over de synaptische spleet, de omkeerbare interactie met de nicotine Ach-receptor en tenslotte de hydrolyse door AChE, vindt plaats binnen enkele milliseconden. Het gehele proces is zowel ruimtelijk als temporeel nauw geïntegreerd (40). Overeenkomstig zijn klassieke rol bedraagt zijn omlooptijd 150 usec, hetgeen overeenkomt met de hydrolyse van 5000 Ach-moleculen/molecuul enzym/seconde. Het hydrolyseert het substraat zo snel dat de concentratie rond het enzymmolecuul uitgeput raakt ten opzichte van de concentratie in de bulkoplossing.
Al meer dan twintig jaar wordt verondersteld dat AChE naast zijn klassieke rol in het beëindigen van synaptische transmissie nog andere rollen vervult. Dergelijke niet-klassieke functies van AChE zouden de hydrolyse van Ach in een niet-synaptische context kunnen inhouden. Van het 26 residuen tellende C-eind peptide ARP (acetylcholinesterase read through peptide) van het menselijke AChE, dat wordt geproduceerd wanneer de R splice variant door stress wordt geïnduceerd, is bekend dat het de hematopoïsche differentiatie moduleert (41).
Verschillende in vivo en in vitero studies in het centrale zenuwstelsel suggereren dat ontwikkelingsgereguleerd AChE enzym een rol speelt in niet cholinerge functies zoals morfometrische processen, celdifferentiatie en synaptogenese langs het zenuwstelsel (20).Er zijn tal van studies die suggereren dat een van de niet klassieke rollen van AChE zou kunnen zijn als een adhesie-eiwit betrokken bij synaptische ontwikkeling en onderhoud (42). Bepaalde andere bewijzen impliceren AChE als botmatrix eiwit, en onlangs is aangetoond dat het interageert met het baseleiwit laminine (43, 44).
Er zijn verschillende studies die hebben aangetoond dat AChE betrokken is bij de groei van neurieten. Vormen van AChE die Ach hydrolyseren maar de corboxy-terminale (verantwoordelijk voor neuritogene activiteiten), neuritische domeinen misten, slaagden er niet in neuritische groei te bevorderen, hetgeen de onafhankelijkheid van de katylitische en neuritogene activiteit van elkaar aantoont (19). Er is ook voorgesteld dat AChE meerdere rollen speelt in de embryogenese. Zebravissen vormen een uitstekend systeem om de invivo rollen van AChE tijdens embryogenese te onderzoeken. Een missence mutatie in het ache gen van de zebravis werd geïdentificeerd die de hydrolyse van Ach opheft maar de neuritogene corboxy-terminal intact laat (45). In het geval van de ache mutant embryo’s werden ook defecten gerapporteerd in de ontwikkeling van spiervezels en primaire sensorische neuronen overleving en dendritische groei. Deze defecten werden collectief geïnterpreteerd om niet klassieke rollen van AChE te ondersteunen.
Er zijn voldoende bewijzen ter ondersteuning van zijn rol in de hydrolyse van substance P, met, leu-enkefaline en de afbraak van andere neuropeptiden ook (46, 47).
AChE is ook voorgesteld om functie in het hart morfogenese. AChE activiteit is gedetecteerd in embryonale ratten- en kippenharten vóór de innervatie (48, 49). Hoewel de rol van AChE in het embryonale hart onduidelijk is, is voorgesteld om een embryonaal prikkelgeleidingssysteem te reguleren (50). Er zijn echter aanvullende functies nodig om de rol van AChE in de hartfunctie verder te onderzoeken.
AChE heeft aangetoond de assemblage van αβ peptide tot amyloïde fibrillen te versnellen, waarschijnlijk door interactie aan de perifere zijde (51-53).Ook is aangetoond dat AChE / αβ complexen een verhoogde neurotoxiciteit vertonen in vergelijking met fibrillen die alleen αβ bevatten (54).
In een farmacologische context hebben de studies aangetoond dat niet-klassieke activiteiten van AChE lijken te convergeren op de perifere anionische plaats.
Moleculaire diversiteit
Het enzym AChE komt voor in een reeks moleculaire vormen die verschillen zowel in quaternaire structuur als in de wijze van verankering binnen de synaps, gegenereerd door alternate splicing van het C-eind exon van één enkel gen, gevolgd door post translationele modificatie (55, 56). Acetylcholinesterase (AChE) bestaat in zes verschillende vormen (isoenzymen) nl. globulair monomeer (G1), dimeer (G2) en tetramer (G4); tetramer met staart (A4), tetramer met dubbele staart (A8) en tetramer met drievoudige staart (A12) (fig.3). De meest rationele terminologie op basis van deze informatie is die welke door Bon et al. (1982) is ontwikkeld. In hun systeem worden de bolvormige vormen aangeduid met “G” en de staartvormen met “A”.
Monomeer- en disulfide-gekoppelde dimeervormen kunnen oplosbaar zijn of met een glycofosfolipide aan een membraan vastzitten. De tetramervormen kunnen oplosbaar zijn, met lipiden aan een membraan gekoppeld zijn of aan een drievoudige collageenhelix gehecht zijn. Monomere vormen worden over het algemeen opgenomen in synapsen van het centrale zenuwstelsel van gewervelde dieren, terwijl bij de neuromusculaire juncties drie katalytische subeenheden van tetrameren zijn gehecht aan een drievoudige spiraalvormige collageenstaart die vervolgens verankerd is aan de basale lamina binnen de synaptische spleet.
De expressie van AChE moleculaire vormen, in Torpedo, werd waargenomen weefsel-specifiek wat suggereert dat polymorfisme vertoond door AChE afhankelijk is van een aantal factoren waaronder
de hoeveelheid vrijkomende Ach
de grootte van de synaptische ruimte en/of
het temporele verloop van de fysiologische stimulatie.(58)
Regulatie van AChE-expressie in neuronen
In de afgelopen jaren zijn er uitgebreide studies verricht om de moleculaire mechanismen en routes te ontrafelen die betrokken zijn bij AChE-expressie en -lokalisatie in neuronale en niet-neuronale weefsels. AChE komt sterk tot expressie tijdens de neuronale differentiatie. Er is waargenomen dat de AChE – 3′ onvertaalde regio (UTR) een AU rijk element (ARE) bevat dat direct interageert met het RNA bindend eiwit ‘HuD’ wat resulteert in de overvloed van AChE transcripten in neuronale cellen (59). Deze bevindingen tonen het belang aan van post-transcriptionele mechanismen bij het reguleren van AChE expressie in differentiërende neuronen en impliceren ‘HuD’ als een belangrijke trans-werkende factor in deze gebeurtenissen. Daarnaast zijn er echter nog verschillende andere regulerende factoren die de stabiliteit, lokalisatie en translatie van AChE transcripten bepalen. Vele cis- en trans-actoren en eiwitten staan in wisselwerking met AChE – 3′-UTR. (59).
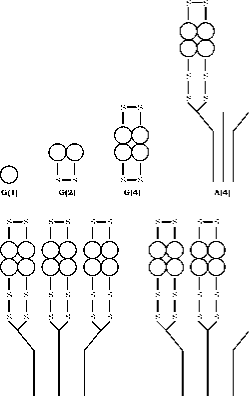
Figuur 3. Kwartaire structuur van de zes hoofdvormen van AChE. Globulaire vormen worden weergegeven door G en asymmetrische vormen door A, volgens het schema van Bon et. al. (16). De plaats van de disulfidebruggen en de wijze van binding aan de collageenstaart zijn schematisch weergegeven volgens het model van Anglister en Silman.(57)
AChE in neurodegeneratieve en neuromusculaire aandoeningen :
Acetylchlinesterase is rechtstreeks in verband gebracht met bepaalde neuromusculaire aandoeningen zoals myasthenia gravis,glaucoom (60),en meest recentelijk met het verlichten van de cholinerge deficiëntie geassocieerd met de ziekte van Alzheimer (61).
Clinisch gezien is matige remming van AChE effectief bij de behandeling van deze ziekten om het effect van ACh op de receptor te verlengen Een dergelijke behandeling is wenselijk als er sprake is van een verlaagde concentratie ACh, zoals bij de ziekte van Alzheimer, of als er minder receptoren zijn, zoals in het geval van myasthenia gravis. Momenteel zijn in de VS door de FDA goedgekeurde remmers voor de behandeling van de ziekte van Alzheimer: E2020 (donepezil, Aricept), tacrine (Cognex), rivastigmine (Exelon) en galantamine (Reminyl)(62). Inhibitoren voor de behandeling van myasthenia gravis omvatten pyridostigmine (Mestinon) en neostigmine (Prostigmine(63). Een overweldigende remming van AChE, met name door covalente binding aan het serine van de actieve plaats, is echter onveranderlijk dodelijk. Daarom is AChE een uitstekend doelwit voor natuurlijk voorkomende toxinen zoals het slangengif fasciculine II, pesticiden zoals parathion en malathion, en chemische strijdmiddelen zoals sarin, tabun en VX (64)
Conclusies
In dit artikel hebben we eerst een overzicht gegeven van de recente resultaten met betrekking tot de biochemie en de moleculaire structuur van AChE, die het unieke enzym de flexibiliteit geeft om in aanraking te komen met diverse legendes en ook om een hoge turn over rate te vertonen. We hebben ook zijn brede histochemische distributiepatroon in neuronale en niet-neuronale weefsels geïllustreerd, wat verder gecorreleerd kan worden met zijn klassieke en niet-klassieke rollen die de revue gepasseerd zijn. Verdere studies over de regulatie van AChE expressie, die het belang van deze post-transcriptionele gebeurtenissen in vivo ontrafelen, zijn van het grootste belang omdat deze uiteindelijk kunnen leiden tot het ontwerpen van aanvullende therapeutische strategieën gericht op het bevorderen van neuronale regeneratie en overleving. De recente ontwikkelingen in de AChE-biologie, met name de implicaties daarvan voor bepaalde neurodegeneratieve aandoeningen, zijn van groot belang en zijn hier kort besproken. Dit onderzoek resulteert in de productie van nieuwe klassen van AChE-remmers gericht tegen zijn PAS (perifere anionische sites), die in de context van de ziekte van Alzheimer een tweeledige rol zouden kunnen spelen door tegelijkertijd de hydrolyse van acetylcholine te remmen en de assemblage van hetá â-peptide tot amyloïde vezel te vertragen.
Afkortingen
AChE – acetylcholinesterase; ARP – acetylcholinesterase read through peptide; Ach – acetycholine; UTR – untranslated region; ARE – AU rich element; PAS – perifere anionische site; Glu – glutaminezuur; His – histidine; Ser – serine; Asp – asparaginezuur; Trp – tryptofaan; Tyr – tyrosine; Phe – fenylalanine.