1. Morfologie
2. Functies
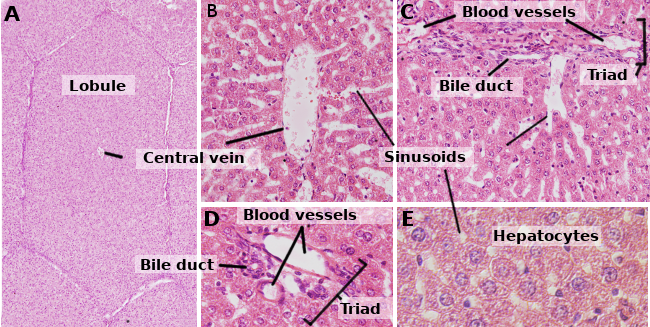
Hepatocyten zijn de cellen van de lever, die ongeveer 80 % van dit orgaan uitmaken. Ze zijn georganiseerd in bladen van ongeveer één cel dik. De vellen zijn onderling verbonden en vormen een sponsachtige structuur (Figuren 1 en 2). Hepatocyten zijn tamelijk langlevende cellen, die zich ongeveer om de 5 maanden vernieuwen. Dit kan echter veranderen onder regeneratieve processen waarbij hepatocyten een groot vermogen tot proliferatie en regeneratie van beschadigd leverweefsel vertonen.

Morfologie
Hepatocyten zijn polyedrale cellen, dat wil zeggen dat ze verschillende gezichten hebben. Meestal hebben ze 6 vlakken, maar het aantal kan variëren. De vlakken staan in contact met hetzij andere hepatocyten, hetzij met een sinusoïde (figuur 3). Hepatocyten zijn grote cellen, ongeveer 20 tot 30 µm in diameter. Zij vertonen een ronde kern gecentreerd in het cytoplasma. In de lever van volwassen mensen kan echter tot 25 % van de hepatocyten binucleair zijn (figuur 2). De meeste kernen zijn tetraploïd, d.w.z. dat zij een dubbele hoeveelheid DNA bevatten dan een gewone cel. De grootte van de kernen is variabel, hoewel zij groter zijn bij tetraploïde cellen. De kernen vertonen verspreide heterochromatine en een of meer nucleoli. Het is niet gebruikelijk om mitotische hepatocyten (1 op de 10000 of 20000 hepatocyten) waar te nemen in normale omstandigheden. Echter, de mitotische hepatocyten nemen enorm toe onder leverletsels en regeneratieprocessen. De kenmerken van het cytoplasma variëren naar gelang van de fysiologische toestand van de cel, vooral onder invloed van vet- en glycogeendepots. Er zijn veel kleine mitochondriën, 800 tot 100 per hepatocyt. Men schat dat één hepatocyt ongeveer 50 Golgi-apparaten kan bevatten die gewoonlijk georganiseerd zijn in stapels van 3 tot 5 cisternen met verdikte laterale gebieden die donkere lichamen bevatten. De stapels cisternen liggen op de een of andere manier verspreid in het cytoplasma, hoewel ze dikwijls dicht bij biliaire canaliculi worden waargenomen (Figuren 3 en 4). Hepatocyten bevatten veel peroxisomen (200 tot 300), meer dan andere regelmatige cellen. In de buurt van de billiaire kanaaltjes worden ook veel lysosomen aangetroffen.


In het cytoplasma van de hepatocyten worden talrijke glycogeen- en lipide-depots aangetroffen (het korrelige uiterlijk van de hepatocyten na hematoxyline- en eosinekleuring is te wijten aan de gaten die zijn achtergebleven door lipide-extractie uit het cytoplasma tijdens de weefselverwerking). In het cytoplasma bevinden zich ook restlichaampjes die lipofuscine bevatten. Het gladde endoplasmatisch reticulum is vrij talrijk, hoewel de grootte van de organellen varieert naar gelang van de metabole activiteit van de hepatocyt. Het is geconcentreerd rond glycogeendepots. In de leverkwabben zijn er morfologische verschillen bij vergelijking van perifere en centrale hepatocyten, die meestal worden beïnvloed door de bloedkenmerken. Zo slaan de perifere hepatocyten na de spijsvertering als eerste glycogeen op, maar zij zijn de laatsten om dit glycogeen weer op te slaan wanneer de rest van het lichaam er om vraagt. Vetopslag gebeurt echter het eerst in de centraal gelegen hepatocyten, die gewoonlijk een gladder endoplasmatisch reticulum hebben. Anderzijds is het ruwe endoplasmatische reticulum 50% groter in de perifeer en gemiddeld gelegen hepatocyten dan in die in het binnenste deel van de leverkwab.
In tegenstelling tot andere epitheelcellen zijn hepatocyten niet gebonden aan een basaal membraan. Hun basolaterale membranen zijn omgeven door een extracellulaire matrix met lage dichtheid, die door de hepatocyten zelf wordt gesynthetiseerd. Deze vergemakkelijkt de diffusie en de uitwisseling van moleculen met de sinusoïden via de ruimte van Disse, of perisinousoïdale ruimten, die de ruimte is tussen het endotheel met venestertjes en de hepatocyten. Deze extracellulaire matrix ontbeert laminine, althans wanneer de hepatocyt gedifferentieerd is. Collageen type IV en fibronectine lijken echter noodzakelijk voor een goede differentiatie van de hepatocyten. Hepatocyten zijn onderling met elkaar verbonden door gap junctions, adherente junctions, desmosomen en tight junctions.
Hepatocyten zijn gepolariseerde cellen, dat wil zeggen dat er verschillen zijn tussen de gebieden die naar de galkanalen gericht zijn en de gebieden die dicht bij de sinusoïden liggen. De polariteit is essentieel voor de correcte functie van de hepatocyt, en is ontregeld in vele lever pathologieën. De apicale regio staat in contact met de galkanaaltjes. Net als in het apicale domein van epitheelcellen zijn er tight junctions, die de integriteit van de galkanalen afdichten en in stand houden. Het apicale membraan vouwt zich in microvilli die het membraanoppervlak enorm vergroten. Het apicale membraan is ongeveer 13% van het totale hepatocytenmembraan, en het kan een grote hoeveelheid moleculen bevatten. Verwijdering van tight junctions leidt tot disorganisatie van de celpolariteit. Hepatocytenpolariteit en galkanalen komen tot stand tijdens de embryonale ontwikkelingsperiode.
De functionele polariteit berust op een ongelijke verdeling van transporters en andere membraanmoleculen tussen de apicale en de baso-laterale plasmamembraandomeinen. ABC transporters (ATP binding cassettes) behoren tot de belangrijkste apicale transporters in hepatocyten. Het Golgi-apparaat, endosomen en het cytoskelet (microtubuli en actinefilamenten) zijn verantwoordelijk voor de differentiële distributie van moleculen tussen de twee membraandomeinen. Er zijn twee aanvoerroutes van eiwitten naar het apicale domein (Figuur 5): vanuit het Golgi-apparaat worden de eiwitten (bijvoorbeeld ABC-transporters) in vesikels afgegeven naar het apicale plasmamembraan of naar recyclerende endosomen, die als intermediair fungeren. Andere eiwitten volgen een transcytose-route, waarbij ze eerst naar de basolaterale membranen reizen en vervolgens worden ingesloten in endocytose-blaasjes naar endosomen, waar ze opnieuw in blaasjes worden verpakt en naar het apicale membraan worden getransporteerd. Zeldzamer is een route die de exocytose van lysosomen inhoudt, gevolgd door coöper transporters.

Functies
De belangrijkste functie van hepatocyten is het metaboliseren van stoffen die afkomstig zijn van de spijsvertering. De lever wordt bevloeid door de poortader die moleculen verzamelt die afkomstig zijn van de spijsvertering in de darm. De hepatocyten zijn ook sterk betrokken bij de ontgifting van potentieel schadelijke moleculen. Anderzijds synthetiseren hepatocyten gal, die uiteindelijk in de darm vrijkomt en helpt bij de vertering. Voor beide functies, het metaboliseren van moleculen uit de spijsvertering en het afgeven van gal, bevinden de hepatocyten zich op een bevoorrechte plaats: in contact met de sinusoïden, die de in de darm verteerde moleculen brengen, en de galkanalen vormen die de gal afvoeren uit de lobben van de lever.
Glucosespiegels. Hepatocyten halen glucosemoleculen uit de spijsvertering en slaan deze op als glycogeen, dat wordt gemobiliseerd wanneer het lichaam energie nodig heeft. Glycogeen wordt meestal aangetroffen in de buurt van het endoplasmatisch reticulum omdat het enzym glucose-6-fosfatase zich in dit organel bevindt. Glucose-6-fosfatase katalyseert glucose-6-fosfaat, de moleculaire vorm van glucose na glycogeenkatabolisme, en produceert vrije glucose, die de hepatocyt kan verlaten en de bloedstroom kan bereiken.
Synthese van moleculen. Galzouten, die helpen bij de vertering van vet, zijn een van de stoffen die door de hepatocyten worden gesynthetiseerd. In het gladde endoplasmatisch reticulum bevinden zich vele enzymen die betrokken zijn bij de synthese van cholesterol en andere lipiden. Bovendien produceren de hepatocyten lipoproteïnen die nodig zijn voor het transport van lipiden in de bloedstroom. Fibrinogeen voor de bloedstolling en plasma-albumines worden eveneens door de hepatocyten gesynthetiseerd. In de lever wordt ureum geproduceerd als bijproduct van de afbraak van eiwitten. De productie en accumulatie van grote hoeveelheden ureum in het organisme kan schadelijk zijn. Hepatocyten slaan vitamine A en B, en heparine op.
Lipidenmetabolisme. Bèta-oxidatie, betrokken bij het vetkatabolisme, is werkzaam in de overvloedige peroxisomen van de hepatocyten.
Detoxificatie. Hepatocyten verzamelen voedzame stoffen die afkomstig zijn van de spijsvertering, maar zij zijn ook de eersten die mogelijk giftige stoffen ontvangen. Ethanol van alcoholische dranken wordt hoofdzakelijk in de lever afgebroken, in feite in de vele peroxisomen van de hepatocyten. In deze organellen wordt de helft van de ingenomen alcohol omgezet in acetaldehyde. In het gladde endoplasmatische reticulum bevinden zich enzymen die betrokken zijn bij de afbraak of inactivering van toxinen en geneesmiddelen. Tijdens perioden waarin veel toxische stoffen moeten worden verwijderd, zoals tijdens medicijnbehandelingen of voortdurend alcohol drinken, kan het endoplasmatisch reticulum het meest omvangrijke organel van de hepatocyt worden. Geneesmiddelen worden gewoonlijk geïnactiveerd door conjugatie met andere moleculen. Bijvoorbeeld, glucosyltransferase conjugeert moleculen met barbituraten.
Het opslaan en reguleren van ijzer. Hepatocyten kunnen ijzer opslaan, dat geconcentreerd is in cytoplasmatische depots die gebonden zijn aan ferritine. Hepatocyten kunnen ijzer op verschillende manieren vastleggen: gebonden aan transferrine, als onderdeel van heemgroepen en van niet-heemgroepen. Transferrine-ijzer komt de cel binnen door TRF1-receptor gemedieerde endocytose. Wanneer endocytische blaasjes versmelten met endosomen, komt transferrine Fe3+ vrij, dat wordt omgezet in F2+ en naar het cytosol wordt geëxtrudeerd door DMT1 (divalent metaal transporter 1) transporter. Hemen-ijzermoleculen worden ook geëndocytiseerd en door het endosomale membraan naar het cytosol getranslokeerd door de HRG1-transporter. Het grootste deel van het ijzer komt echter vanuit de extracellulaire ruimte binnen via de ZIP14-transporter die in het plasmamembraan van de hepatocyt tegenover de sinusoïden is geplaatst. Eenmaal in het cytosol wordt het ijzer gebonden aan ferritine en opgeslagen in het cytoplasma omdat het vrije ijzer giftig is. Het ijzer wordt uit de enterocyt vrijgemaakt door de ferroportine-transporter die zich in het plasmamembraan bij de sinousoïden bevindt.
Na het beenmerg is de lever het tweede belangrijke productiecentrum van heemgroepen. Hemgroep is een prostetische groep (niet peptidisch) die aanwezig is in verschillende eiwitten voor het transport van zuurstof, in enzymen zoals katalasen en peroxidasen die beschermen tegen oxiderende stoffen. Zij maakt ook deel uit van de mitochondriale en peroxysomale cytochromen. De grotere hoeveelheid heemgroepen wordt aangetroffen in de hemoglobine, die in het beenmerg worden gesynthetiseerd. In de lever is de synthese van heemgroepen afhankelijk van de hoeveelheid microsomale p450-cytochromen die de cel nodig heeft, zodat de meeste van deze heemgroepen deel uitmaken van p450-cytochromen.
Hepatocyten geven het hormoon hepcidine af, dat de systemische ijzerconcentratie in het lichaam regelt. Dit hormoon regelt de hoeveelheid ijzer in het plasma door de internalisering en afbraak te bevorderen van ferroportine, een ijzertransporter die wordt aangetroffen in enterocyten, macrofagen en hapatocyten. Verwijdering van ferroportine remt het vrijkomen van ijzer uit deze cellen. De hepcidinesynthese wordt gereguleerd door de transferrine-ijzerconcentratie in het plasma, door ijzerdepots in de hepatocyten en door ontstekingen. De erytropoëtische activiteit remt de afgifte van hepcidine.
Bibliografie
Gissen P, Arias IM. 2015. Structurele en functionele hepatocytenpolariteit en leverziekten. Tijdschrift voor hepatholotie. 63: 1023-1037. ![]()
Knutson MD. 2014. Adult intestinal stam cells: critical drivers of epithelial homeostasis and regeneration. Nature review in moleculaire celbiologie. 15:19-33.
Weiss L, Greep RO. 1982. Histología. 4ªedición. Editorial el Ateneo. Barcelona.