Annals of Neurosciences, Volume 15, Issue 4 (October), 2008
Anurag Tripathi1, UC Srivastava2
1Department of Zoology, Ranchi College,(Ranchi University) Ranchi
2Department of Zoology, University of Allahabad, Allahabad, India – 211 002
Corrosponding Author
Dr UC Srivastava
Professor of Zoology
Department of Zoology, University of Allahabad
Allahabad, India – 211 002
Phone : 93351 20178
E-mail : [email protected]
(Data otrzymania: 08.09.2008)
Abstract
Acetylocholinesteraza (AChE) kończy neurotransmisję w synapsach cholinergicznych poprzez rozszczepienie neuroprzekaźnika acetylocholiny. Natura i rozmieszczenie enzymu zostały dokładnie zbadane u wielu bezkręgowców i kręgowców, w tym u ludzi, zarówno pod względem histochemicznym, jak i biochemicznym. Enzym wykazuje wysoki stopień zmienności w rozmieszczeniu, z godną uwagi obecnością w tkankach nieneuronalnych, co dostarcza istotnych tematów do badania jego nieklasycznej roli.
Ostatnio pojawiło się wiele informacji dotyczących jego dynamicznej struktury, ekspresji genów, roli w morfogenezie neuronów i synaptogenezie. Znaczenie AChE wynika z faktu, że jest ona celem leków przeznaczonych do leczenia miastenii gravis, jaskry, choroby Alzheimera i in.
Pamiętając o powyższych faktach, w niniejszym artykule dokonano dokładnego przeglądu jej biochemii, dynamiki strukturalnej, szerokiej dystrybucji, izoform i ich implikacji w zaburzeniach neurodegeneracyjnych.
Słowa kluczowe : Acetylocholinoesteraza, Cholinergiczna, Formy molekularne, Zaburzenia neurodegeneracyjne, Choroba Alzheimera.
Wprowadzenie
Acetylocholinoesteraza ( AChE) jest jednym z najbardziej wydajnych enzymów układu nerwowego, który koncentruje się w synapsach cholinergicznych oraz w synapsach nerwowo-mięśniowych, gdzie szybko hydrolizuje neuroprzekaźnik acetylocholinę (Ach) do choliny i octanu, odgrywając tym samym zasadniczą rolę w neurotransmisji cholinergicznej.
Termin acetylocholinoesteraza został wprowadzony w 1949 roku przez Augustintiona i Nachmansohna dla specyficznej cholinoesterazy zdolnej do hydrolizy acetylocholiny szybciej niż inne esterazy. W 1964 roku Komisja Enzymologiczna zarekomendowała nazwę „Acetylocholinesteraza” (Acetylcholine Acetyl hydrolase; 3.1.1.7) dla prawdziwej i specyficznej cholinoesterazy.
Rozmieszczenie enzymu w tkankach nerwów centralnych i obwodowych różnych kręgowców wykazuje duży zakres zmienności (1-13). Odnotowano jego lokalizację w tkankach innych niż neuronalne oraz w komórkach glejowych (14, 15). Enzym wykazuje również różnorodność molekularną z jego sześcioma różnymi formami molekularnymi i dynamiką strukturalną, co ułatwia jego powinowactwo i działanie z różnymi legendami (16-17). Ponadto uważa się, że AChE odgrywa kilka nieklasycznych ról, niezależnych od jego funkcji katalitycznej, tj. hydrolizy Ach. Te klasyczne i nieklasyczne role AChE ilustrują adekwatność jej szerokiego występowania w tkankach neuronalnych i nieneuronalnych (18-20).
Znaczenie AChE w homeostazie organizmu jest podkreślone przez fakt, że są one celem niektórych z najsilniejszych toksyn, w tym środków owadobójczych, jadu węża i broni chemicznej (21).
Dlatego tak szeroka dystrybucja i różne funkcje, formy molekularne, dynamika strukturalna itp. AChE zapewniają odpowiednią podstawę do uznania go za wszechstronny enzym, którego szczegółowa znajomość pomoże w projektowaniu specyficznych leków zwalczających różne choroby neurodegeneracyjne związane z tym enzymem.
Biochemia i dynamika strukturalna
Trójwymiarowa struktura AChE została po raz pierwszy określona przez Joela Sussmana i wsp. w 1991 roku, przy użyciu białka z T. californica (22). Struktury domen katalitycznych AChE z gatunków takich jak T. californica, mysz i człowiek są dość podobne (rys.1) (23, 24).
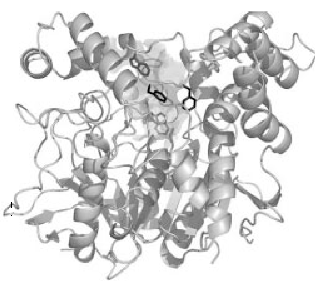
Rysunek 1. Trójwymiarowy diagram wstęgowy acetylocholinoesterazy Torpedo Californica.
Acetylocholinoesteraza jest hydrolazą serynową należącą do rodziny korboksylastrów typu B w obrębie wyższych eukariotów. Jest to elipsoidalna cząsteczka o wymiarach około 45×60×65 A0, która składa się z 12-nitkowego mieszanego arkusza beta otoczonego 14 helikami alfa. (22). W stanie naturalnym jest monomerem o masie cząsteczkowej około 60 000 i tworzy agregaty, które w dalszym ciągu wytwarzają aktywność katalityczną.
Miejsce aktywne składa się z dwóch podstref, podstrefy estrowej posiadającej triadę katalityczną (Ser-His-Glu) oraz peryferyjnej wiążącej podstrefy anionowej (PAS), która mieści dodatni biegun czwartorzędowy Ach. ( Rys. 2a)
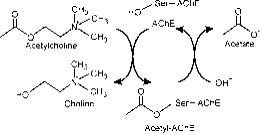
Figura 2a: Różne miejsca działania AChE; miejsce katalityczne znajduje się w głębokim wąwozie.
Podstrykt estrowy zawiera maszynerię katalityczną enzymu, triadę Ser 200, His-440 i Glu-327 (liczby oznaczają sekwencję aminokwasów w łańcuchu polipeptydowym). Ta triada jest podobna do innych proteaz serynowych, z wyjątkiem tego, że triada ta po raz pierwszy wykazuje Glu jako trzeciego członka, w przeciwieństwie do Asp. Ponadto triada ta jest przeciwnie skierowana niż w przypadku innych proteaz. Podstrykt anionowy definiowany jest przez Trp 84, Phe 330, Phe 331. Jej rolą jest orientowanie naładowanej części substratu, który wchodzi do miejsca aktywnego. Realizowane jest to głównie przez Trp. 84 (22).
Wąwóz aromatyczny w białku ma głębokość około 20 angstremów i przenika przez połowę drogi enzymu. Miejsce aktywne leży u podstawy tego wgłębienia tylko 4 angstromy powyżej podstawy. Wąwóz aromatyczny jest bardziej odpowiednim określeniem, ponieważ 40% jego wyściółki składa się z 14 reszt aromatycznych, które są wysoce konserwowane w różnych gatunkach AChE (25).
Perferyjne miejsce anionowe ma zdolność wiązania się z wieloma różnymi typami ligandów, przez co wpływa na potwierdzenie miejsca aktywnego. Sześć reszt stabilizuje aktywność w obrębie miejsca aktywnego. Trp 286, Ty r 72, Tyr 124, Glu 285, Asp 74, Tyr 341 zlokalizowane na przeciwległej stronie wejścia do wąwozu. Taki układ reszt zapewnia elastyczność, która umożliwia przyjęcie wielu ligandów, a także implikuje ich ruchliwość konfirmacyjną. AChE katalizuje hydrolizę AChE w następujący sposób (Ryc. 2b):
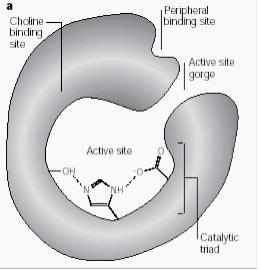
Ryc. 2b: Reakcja substratowa enzymu ilustrująca hydrolizę acetylocholiny.
Dynamika molekularna AChE jest utrzymywana poprzez mechanizm cross talk, w którym interakcje pomiędzy dwoma podstryktami zachodzą w szczególności pomiędzy resztami Trp 286, Trp 86. Kiedy Trp 286 jest związany na peryferiach, wpływa to na Trp 86 w miejscu aktywnym i powoduje odrębne miejsce potwierdzenia. Asp 74 również odgrywa rolę w modulacji allosterycznej enzymu. Wrażliwość tych reszt i plastyczność miejsca aktywnego są prawdopodobnie wynikiem ewolucyjnego projektu mającego na celu nadanie optymalnej aktywności w szerokiej gamie warunków (26.)
Występowanie
Enzym AChE jest obecny w wysokim stężeniu we wszystkich typach tkanek przewodzących, nerwowych i mięśniowych, tkankach centralnych i obwodowych, włóknach ruchowych i czuciowych, współczulnych i przywspółczulnych tzw. cholinergicznych i niecholinergicznych oraz we wszystkich regionach, gdzie znajdują się ciała i połączenia komórkowe, stężenie enzymu jest wysokie(27). Metodami histochemicznymi AChE wykazuje się na ogół w strukturach neuronalnych: perikaryi, błonach powierzchniowych, aksonach, dendrytach i regionach synaptycznych 4, 15, 28-30). Aktywność AChE jest wyższa w neuronach ruchowych niż w neuronach czuciowych (4, 15, 31). Ogólnie uznaje się, że acetylocholinoesteraza w tkankach nerwowych reprezentuje neurony, które zawierają i przypuszczalnie uwalniają acetylocholinę jako neuroprzekaźnik (32).
Wcześniejsze badania miejsca syntezy AChE w obrębie tkanki nerwowej wskazywały na pochodzenie z ciała komórkowego neuronu (33-35). Stwierdzono, że enzym jest związany z retikulum endoplazmatycznym ciała komórkowego i aparatem Golgiego (8, 36). AChE była również związana we frakcjach subkomórkowych mózgu z frakcjami mikrosomalnymi i surowymi frakcjami mitochondrialnymi (2, 37). Dalsze frakcjonowanie surowej frakcji mitochondrialnej z tkanki mózgowej wykazało wysoki poziom aktywności AChE we frakcji błony synaptosomalnej (6).
Większość AChE została zlokalizowana w aksonach i związana z błoną aksonalną (9). Aktywność cholinoesterazy ograniczona do aksonów, została zaobserwowana na pęcherzykach aksonalnych wszystkich nie mielinowanych i niektórych mielinowanych włókien nerwowych (38). Mielinowane włókna nerwowe mają generalnie niższe stężenie niż tzw. niemielinowane, a istota biała zawiera znacznie mniej AChE niż istota szara (27). W izolowanych motoneuronach aktywność AChE jest około dziesięciokrotnie większa w cytoplazmie, dendrytach i aksonach niż w nukleoplazmie, a w nukleolusie nie występuje żadna aktywność (39). Wykazano, że komórki glejowe zawierają AChE w różnych stężeniach (14, 15)
Role klasyczne i nieklasyczne
Cały przebieg przekaźnictwa cholinergicznego, szczególnie na złączu nerwowo-mięśniowym, a mianowicie uwolnienie Ach, jej dyfuzja przez szczelinę synaptyczną, odwracalna interakcja z nikotynowym receptorem Ach i wreszcie hydroliza przez AChE zachodzi w ciągu kilku milisekund. Cały ten proces jest ściśle zintegrowany przestrzennie i czasowo (40). Zgodnie z klasyczną rolą AChE, jego czas rotacji wynosi 150 usec, co odpowiada hydrolizie 5000 cząsteczek Ach/molekułę enzymu/sek. Hydrolizuje on substrat tak szybko, że stężenie wokół cząsteczki enzymu ulega zubożeniu w stosunku do jego stężenia w roztworze objętościowym.
Od ponad dwudziestu lat przypuszcza się, że AChE, oprócz swojej klasycznej roli w przerywaniu transmisji synaptycznej, pełni także inne role. Takie nieklasyczne funkcje AChE mog± być zwi±zane z hydroliz± Ach w kontek¶cie innym niż synaptyczny. 26 reszt C-końcowego peptydu ARP (acetylocholinoesterase read through peptide) ludzkiej AChE, który jest wytwarzany, gdy wariant splice R jest indukowany przez stres, został zgłoszony do modulowania różnicowania hematopoitów (41).
Różne badania in vivo i in vitero w ośrodkowym układzie nerwowym sugerowały, że regulowany rozwojowo enzym AChE odgrywa rolę w nie cholinergicznej funkcji jak procesy morfometryczne, różnicowanie komórek i synaptogeneza wzdłuż układu nerwowego (20).Istnieje wiele badań, które sugerują, że jedna z nie klasycznych ról AChE może być jako białko adhezyjne zaangażowane w rozwój i utrzymanie synaps (42). Niektóre inne dowody wskazują, że AChE jest białkiem macierzy kostnej, a ostatnio wykazano, że oddziałuje z białkiem podstawnym lamininą (43, 44).
Wiele badań dowiodło, że AChE jest zaangażowana we wzrost neurytów. Formy AChE, które hydrolizują Ach, ale pozbawione są domen neurytogennych (odpowiedzialnych za aktywność neurytogenną), nie zwiększyły wzrostu neurytów, wykazując niezależność aktywności katalitycznej i neurytogennej od siebie (19). Zaproponowano również, że AChE odgrywa wiele ról w embriogenezie. Ryby zebra stanowią doskonały system do badania inwazyjnych ról AChE podczas embriogenezy. Zidentyfikowano mutację missence w genie ache ryb zebry, która znosi hydrolizę Ach, ale pozostawia neurytogenny korboksy-końcowy nienaruszony (45). W przypadku zmutowanych embrionów ache odnotowano również defekty w rozwoju włókien mięśniowych oraz w przeżyciu pierwotnych neuronów czuciowych i wzroście dendrytów. Defekty te zostały wspólnie zinterpretowane jako wspierające nieklasyczne role AChE.
Istnieją liczne dowody na poparcie jej roli w hydrolizie substancji P, met, leu-enkefaliny i degradacji innych neuropeptydów, jak również (46, 47).
AChE została również zaproponowana do funkcji w morfogenezie serca. Aktywność AChE jest wykrywana w embrionalnych sercach szczurów i kurcząt przed unerwieniem (48, 49). Chociaż rola AChE w sercu embrionalnym jest niejasna, zaproponowano, że AChE reguluje embrionalny system przewodzenia impulsów (50). Jednakże dodatkowe funkcje są wymagane do dalszego zbadania roli AChE w funkcji serca.
AChE wykazano, że przyspiesza składanie peptydu αβ w fibryle amyloidowe prawdopodobnie poprzez interakcję w miejscu obwodowym (51-53).Wykazano również, że kompleksy AChE / αβ wykazują zwiększoną neurotoksyczność w porównaniu z włóknami zawierającymi tylko αβ (54).
W kontekście farmakologicznym badania ujawniły, że nieklasyczne działania AChE wydają się zbiegać na obwodowym miejscu anionowym. Można zatem przypuszczać, że w przyszłości powstaną nowe kategorie inhibitorów AChE.
Różnorodność molekularna
Ezzym AChE występuje w szeregu form molekularnych różniących się zarówno strukturą czwartorzędową, jak i sposobem zakotwiczenia w synapsie powstałej w wyniku naprzemiennego splicingu C-końcowego eksonu pojedynczego genu, a następnie modyfikacji posttranslacyjnej (55, 56). Acetylocholinoesteraza (AChE) występuje w sześciu różnych formach (izoenzymach): globularnego monomeru (G1), dimeru (G2) i tetrameru (G4); tetrameru z ogonkami (A4), tetrameru z podwójnymi ogonkami (A8) i tetrameru z potrójnymi ogonkami (A12) (ryc. 3). Najbardziej racjonalną terminologią opartą na tych informacjach jest ta opracowana przez Bon i wsp. (1982). W ich systemie formy globularne oznaczane są przez „G”, a formy ogoniaste przez „A”.
Formy monomeru i dimeru połączone disulfidowo mogą być rozpuszczalne lub przyłączone do błony przez glikofosfolipid. Formy tetramerowe mogą być rozpuszczalne, związane z błoną lipidową lub przyłączone do potrójnej helisy kolagenu. Formy monomeryczne są generalnie wbudowane w synapsy centralnego układu nerwowego kręgowców, podczas gdy w złączach nerwowo-mięśniowych trzy podjednostki katalityczne tetramerów są przyłączone do potrójnej helisy kolagenowej, która zakotwicza je do blaszki podstawowej w szczelinie synaptycznej.
Wyrażenie form molekularnych AChE u Torpedo było specyficzne tkankowo, co sugeruje, że polimorfizm wykazywany przez AChE jest zależny od wielu czynników, w tym
ilości uwolnionej Ach
rozmiaru przestrzeni synaptycznej i/lub
czasowego przebiegu stymulacji fizjologicznej.(58)
Regulacja ekspresji AChE w neuronach
W ostatnich latach prowadzono szeroko zakrojone badania mające na celu odkrycie molekularnych mechanizmów i szlaków związanych z ekspresją i lokalizacją AChE w tkankach neuronalnych i nieneuronalnych. AChE ulega wysokiej ekspresji podczas różnicowania neuronów. Zaobserwowano, że AChE – 3′ region nieulegający translacji (UTR) zawiera element bogaty w AU (ARE), który oddziałuje bezpośrednio z białkiem wiążącym RNA „HuD”, powodując obfitość transkryptów AChE w komórkach neuronalnych (59). Wyniki te wskazują na znaczenie mechanizmów post transkrypcyjnych w regulacji ekspresji AChE w różnicujących się neuronach i wskazują na 'HuD' jako kluczowy czynnik transkrypcyjny w tych wydarzeniach. Jednak stabilnością, lokalizacją i translacją transkryptów AChE rządzi dodatkowo kilka innych czynników regulacyjnych. Wiele elementów działających cis i czynników działających trans oraz białek oddziałuje z 3′-UTR AChE. (59).
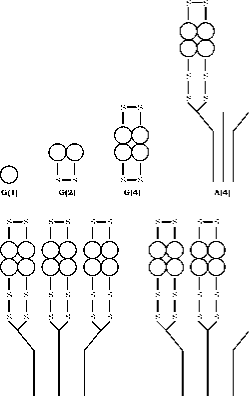
Rysunek 3. Struktura czwartorzędowa sześciu głównych form AChE. Formy globularne są reprezentowane przez G, a formy asymetryczne przez A zgodnie ze schematem Bon et. al. (16). Lokalizacja mostków disiarczkowych i sposób wiązania do ogona kolagenu są przedstawione według modelu skonstruowanego przez Anglistera i Silmana.(57)
AChE w zaburzeniach neurodegeneracyjnych i nerwowo-mięśniowych :
Acetylochlinesteraza jest bezpośrednio związana z niektórymi zaburzeniami nerwowo-mięśniowymi, takimi jak miastenia gravis, jaskra (60), a ostatnio z łagodzeniem niedoboru cholinergicznego związanego z chorobą Alzheimera (61).
Klinicznie, umiarkowane hamowanie AChE jest skuteczne w leczeniu tych chorób w celu przedłużenia działania ACh na receptor Takie leczenie jest pożądane albo w przypadku zmniejszonego stężenia ACh, jak w przypadku choroby Alzheimera, albo w przypadku mniejszej liczby receptorów, jak w przypadku myasthenia gravis. Obecnie zatwierdzonymi przez amerykańską Agencję ds. Żywności i Leków (FDA) inhibitorami w leczeniu choroby Alzheimera są E2020 (donepezil, Aricept), takryna (Cognex), rywastygmina (Exelon) i galantamina (Reminyl)(62). Inhibitory stosowane w leczeniu miastenii gravis obejmują pirydostygminę (Mestinon) i neostygminę (Prostigmine (63). Jednak nadmierne hamowanie AChE, szczególnie przez kowalencyjne wiązanie z seryną w miejscu aktywnym, jest niezmiennie śmiertelne. Stąd, AChE jest głównym celem dla naturalnie występujących toksyn, takich jak fascykulina II z jadu węża, pestycydów takich jak paration i malation oraz chemicznych środków bojowych takich jak sarin, tabun i VX (64)
Wnioski
W niniejszym artykule, po raz pierwszy przeanalizowaliśmy ostatnie wyniki związane z biochemią i strukturą molekularną AChE, która zapewnia unikalność enzymu, elastyczność w napotykaniu różnorodnych legend, a także wykazuje wysoki współczynnik rotacji. Zilustrowaliśmy również jego szeroki histochemiczny wzór dystrybucji w tkankach neuronalnych i nieneuronalnych, który może być dalej skorelowany z jego klasycznymi i nieklasycznymi rolami, które zostały przejrzane. Dalsze badania nad regulacj± ekspresji AChE, które ujawniaj± znaczenie tych post transkrypcyjnych wydarzeń in vivo s± niezwykle istotne, ponieważ ostatecznie mog± one doprowadzić do zaprojektowania dodatkowych strategii terapeutycznych maj±cych na celu promowanie regeneracji i przeżycia neuronów. Ostatnie postępy w biologii AChE szczególnie jego implikacje w niektórych zaburzeniach neurodegeneracyjnych są kluczowym interesem, który został krótko przejrzany. Badania te prowadzą do powstania nowych klas inhibitorów AChE skierowanych przeciwko jej PAS (peryferyjnym miejscom anionowym), które mogą odgrywać podwójną rolę w kontekście choroby Alzheimera poprzez jednoczesne hamowanie hydrolizy acetylocholiny i opóźnianie montażu peptydu amyloidowego.
Skróty
AChE – acetylocholinoesteraza; ARP – acetylocholinoesteraza read through peptide; Ach – acetylocholina; UTR – region nieulegający translacji; ARE – AU rich element; PAS – peryferyjne miejsce anionowe; Glu – kwas glutaminowy; His – histydyna; Ser – seryna; Asp – kwas asparaginowy; Trp – tryptofan; Tyr – tyrozyna; Phe – fenyloalanina.